Difference between revisions of "Lecture 02"
Jump to navigation
Jump to search













































| Line 69: | Line 69: | ||
<tt> H = A C T (not '''G''')</tt><br> | <tt> H = A C T (not '''G''')</tt><br> | ||
<tt> V = G C A (not '''T''')</tt><br> | <tt> V = G C A (not '''T''')</tt><br> | ||
| − | <tt> N = A G C T (a'''n'''y) | + | <tt> N = A G C T (a'''n'''y)</tt> |
]] | ]] | ||
| + | |||
======Slide 009====== | ======Slide 009====== | ||
[[Image:L02_s009.jpg|frame|none|Lecture 02, Slide 009<br> | [[Image:L02_s009.jpg|frame|none|Lecture 02, Slide 009<br> | ||
Revision as of 13:50, 19 September 2007
Update Warning! This page has not been revised yet for the 2007 Fall term. Some of the slides may be reused, but please consider the page as a whole out of date as long as this warning appears here.
(Previous lecture) ... (Next lecture)
The Sequence Abstraction
...
Add:
- Exercises
- Summary points
Lecture Slides
Slide 001

Lecture 02, Slide 001
Slide 002

Lecture 02, Slide 002
Deconstruct this statement: first, we need to consider what it means to compute. Then we need to consider the best representations.
Deconstruct this statement: first, we need to consider what it means to compute. Then we need to consider the best representations.
Slide 003

Lecture 02, Slide 003
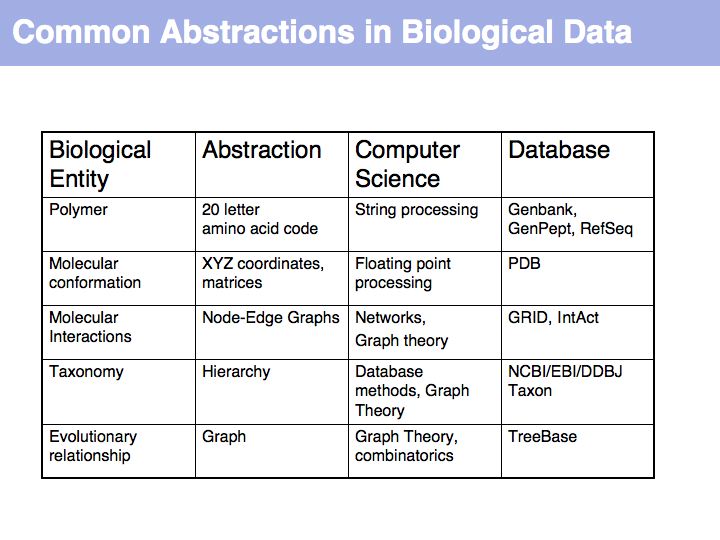
In order to make biology computable, we have to clarify the concepts we are working with. This is useful even beyond the requirements of bioinformatics. It is an exercise in clarifying the conceptual foundations of biology itself. In many instances, definitions in current, common use are deficient, either because our current state of knowledge has gone beyond the original ideas we were trying to subsume with a term (e.g. gene, or pathway), or because an inconsistent formal and colloquial meaning of terms lead to ambiguities (e.g. function), or because the technical meaning of terms is poorly understood and generally misused (e.g. homology).
In order to make biology computable, we have to clarify the concepts we are working with. This is useful even beyond the requirements of bioinformatics. It is an exercise in clarifying the conceptual foundations of biology itself. In many instances, definitions in current, common use are deficient, either because our current state of knowledge has gone beyond the original ideas we were trying to subsume with a term (e.g. gene, or pathway), or because an inconsistent formal and colloquial meaning of terms lead to ambiguities (e.g. function), or because the technical meaning of terms is poorly understood and generally misused (e.g. homology).
Slide 004

Lecture 02, Slide 004
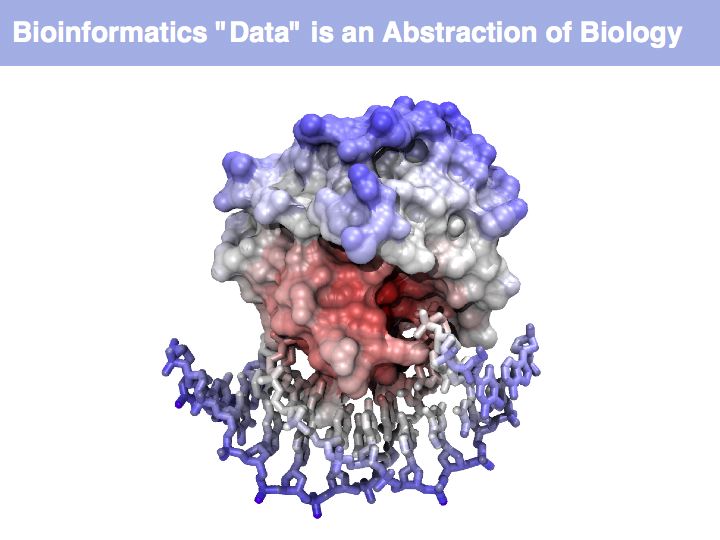
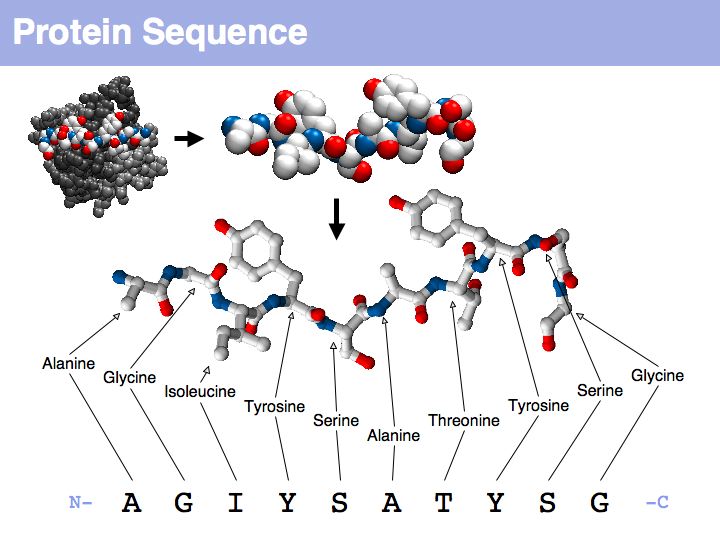
Think about the following question: this image representis a particular biomolecule. What is the best abstraction ?
Think about the following question: this image representis a particular biomolecule. What is the best abstraction ?
Slide 005

Lecture 02, Slide 005
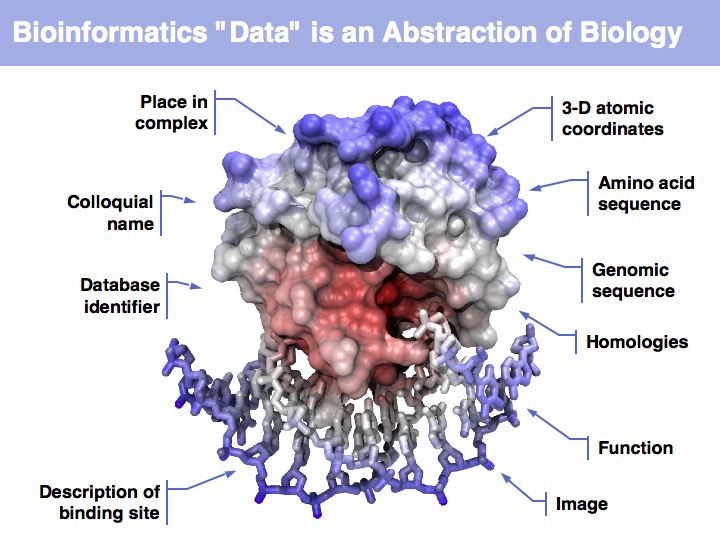
Asking about the best is an ill-posed question if the purpose is not specified. The best for what? There are many possible abstractions, each serving different purposes. Each of the abstractions above can serve as a representation of the biomolecule, each one emphasizes a different perspective and can serve a different purpose.
Asking about the best is an ill-posed question if the purpose is not specified. The best for what? There are many possible abstractions, each serving different purposes. Each of the abstractions above can serve as a representation of the biomolecule, each one emphasizes a different perspective and can serve a different purpose.
Slide 006

Lecture 02, Slide 006
Working with abstractions implies we are no longer manipulating the biological entity, but it's representation. This distinction becomes crucial, when we start computing with representations to infer facts about the original entities! Inferences must be related back to biology! Common problems include that the abstraction may not be rich enough to capture the property we are investigating (e.g. one-letter sequence codes cannot represent amino acid modifications or sequence numbers), or the abstraction may be ambiguous (e.g. one protein may have more than one homologue in a related organism, thus the relationship between gene IDs is ambiguous) or the abstraction may not be unique (e.g. one protein may have more than one function, the same protein name may refer to unrelated proteins in different species).
Working with abstractions implies we are no longer manipulating the biological entity, but it's representation. This distinction becomes crucial, when we start computing with representations to infer facts about the original entities! Inferences must be related back to biology! Common problems include that the abstraction may not be rich enough to capture the property we are investigating (e.g. one-letter sequence codes cannot represent amino acid modifications or sequence numbers), or the abstraction may be ambiguous (e.g. one protein may have more than one homologue in a related organism, thus the relationship between gene IDs is ambiguous) or the abstraction may not be unique (e.g. one protein may have more than one function, the same protein name may refer to unrelated proteins in different species).
Slide 007

Lecture 02, Slide 007
Slide 008

Lecture 02, Slide 008
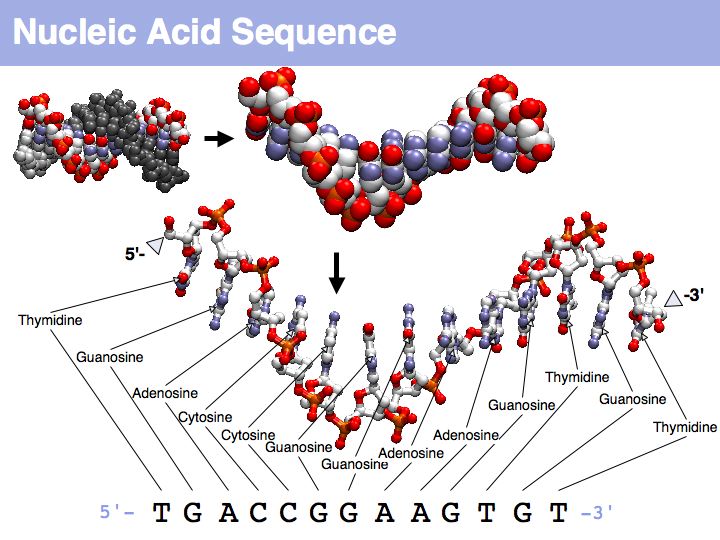
Nucleic acids can form heterocopolymers as DNA or RNA, thus their structural formula can be described (to a close approximation) simply by listing the nucleotide bases in a defined order. A one-letter code has been defined as a shorthand notation for this. By convention, DNA or RNA sequence is written in the direction from the bond with the 5' carbon of the ribose (or deoxyribose), to the bond with the 3'-carbon. Since the 5'-carbon carries a free 5'-phosphate at the terminus of the polynucleotide and the 3'-carbon carries a free OH, this direction happens to be the same direction that nucleic acid polymers are replicated by the DNA-polymerase: a phosphate of a single nucleotide is attached to the free -OH of the polynucleotide. This direction is also the direction of transcription of the RNA polymerase (for the same reason) and (incidentally) the direction of translation by the ribosome. Due to base-pair complementarity, only one strand of a double stranded sequence needs to be recorded since the sequence of the complementary strand is implied: it is simply the reverse complement. The one letter abbreviations are defined by the International Union of Pure and Applied Chemistry (IUPAC) as follows:
A = adenine
C = cytosine
G = guanine
T = thymine
R = G A (purine)
Y = T C (pyrimidine)
K = G T (keto)
M = A C (amino)
S = G C (strong bonds)
W = A T (weak bonds)
B = G T C (not A)
D = G A T (not C)
H = A C T (not G)
V = G C A (not T)
N = A G C T (any)
Nucleic acids can form heterocopolymers as DNA or RNA, thus their structural formula can be described (to a close approximation) simply by listing the nucleotide bases in a defined order. A one-letter code has been defined as a shorthand notation for this. By convention, DNA or RNA sequence is written in the direction from the bond with the 5' carbon of the ribose (or deoxyribose), to the bond with the 3'-carbon. Since the 5'-carbon carries a free 5'-phosphate at the terminus of the polynucleotide and the 3'-carbon carries a free OH, this direction happens to be the same direction that nucleic acid polymers are replicated by the DNA-polymerase: a phosphate of a single nucleotide is attached to the free -OH of the polynucleotide. This direction is also the direction of transcription of the RNA polymerase (for the same reason) and (incidentally) the direction of translation by the ribosome. Due to base-pair complementarity, only one strand of a double stranded sequence needs to be recorded since the sequence of the complementary strand is implied: it is simply the reverse complement. The one letter abbreviations are defined by the International Union of Pure and Applied Chemistry (IUPAC) as follows:
A = adenine
C = cytosine
G = guanine
T = thymine
R = G A (purine)
Y = T C (pyrimidine)
K = G T (keto)
M = A C (amino)
S = G C (strong bonds)
W = A T (weak bonds)
B = G T C (not A)
D = G A T (not C)
H = A C T (not G)
V = G C A (not T)
N = A G C T (any)
Slide 009

Lecture 02, Slide 009
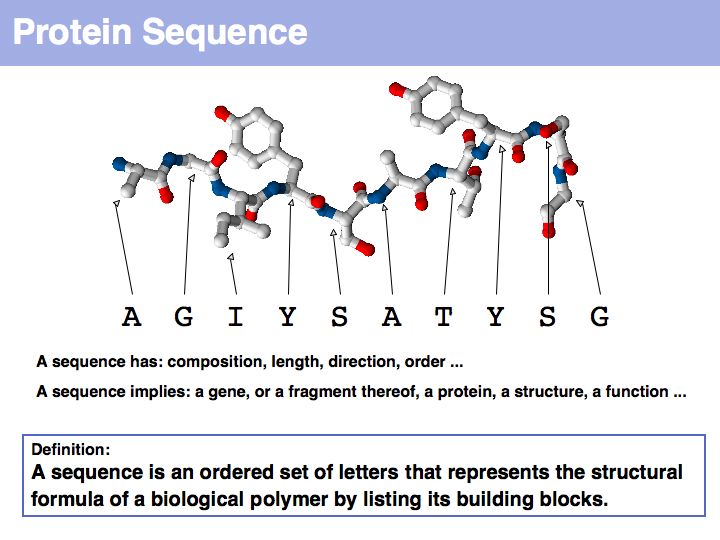
Proteins are amino acid heterocopolymers, thus their structural formula can be described (to a close approximation) simply by listing their constituent amino acid residues in a defined order. The one-letter code is a shorthand notation for this. By convention, protein sequence is written from aminoterminus (N-) to carboxyterminus (C-). This happens to be the same direction in which the protein is synthesized on the ribosome.
Proteins are amino acid heterocopolymers, thus their structural formula can be described (to a close approximation) simply by listing their constituent amino acid residues in a defined order. The one-letter code is a shorthand notation for this. By convention, protein sequence is written from aminoterminus (N-) to carboxyterminus (C-). This happens to be the same direction in which the protein is synthesized on the ribosome.
Slide 010

Lecture 02, Slide 010
Slide 011

Lecture 02, Slide 011
Slide 012

Lecture 02, Slide 012
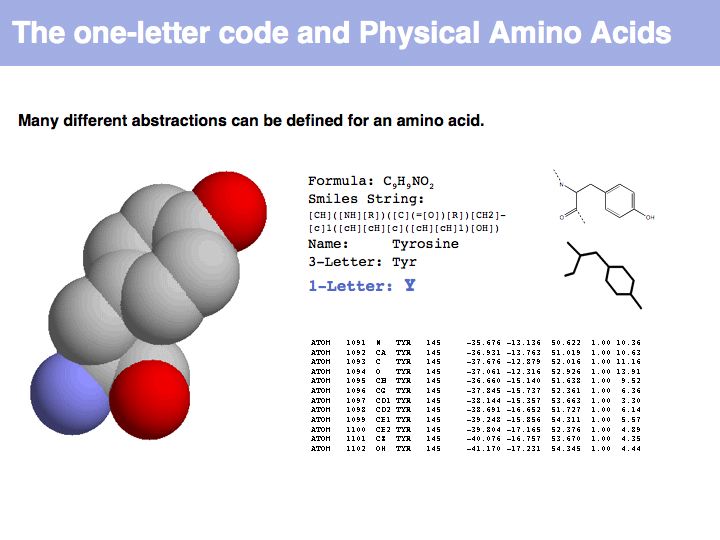
An amino acid is a molecule. A number of abstractions are in common use for this molecule: its chemical formula simply describes the elemental composition, a so called SMILES string (see also here) captures bonding topology as well, its information is equivalent to a chemical graph. A set of records of 3D coordinates can describe the three-dimensional conformation, this can in turn be displayed in a number of different image options - like a simple line drawing or a set of spheres, color coded by element, with relative sizes corresponding to the elements' Van der Waals radii.
An amino acid is a molecule. A number of abstractions are in common use for this molecule: its chemical formula simply describes the elemental composition, a so called SMILES string (see also here) captures bonding topology as well, its information is equivalent to a chemical graph. A set of records of 3D coordinates can describe the three-dimensional conformation, this can in turn be displayed in a number of different image options - like a simple line drawing or a set of spheres, color coded by element, with relative sizes corresponding to the elements' Van der Waals radii.
Slide 013

Lecture 02, Slide 013
... see IUPAC
... see IUPAC
Slide 014

Lecture 02, Slide 014
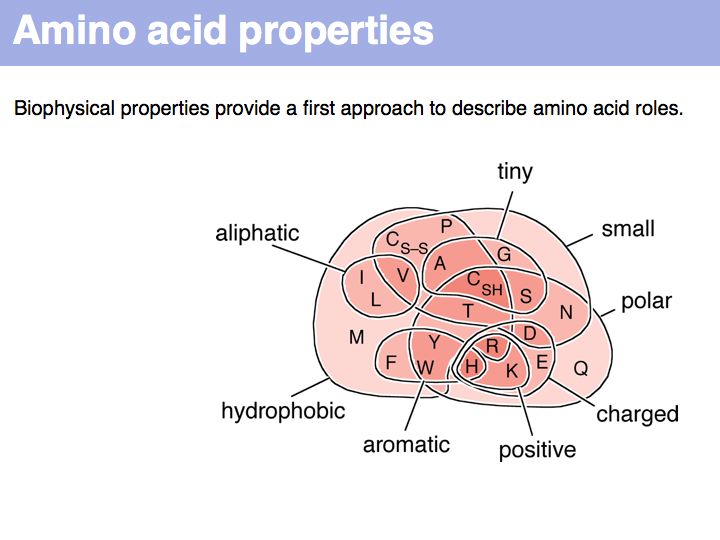
A Venn diagram of biophysical amino acid properties. Of course, the precise role of a paricula amino acid depends on
A Venn diagram of biophysical amino acid properties. Of course, the precise role of a paricula amino acid depends on
Slide 015

Lecture 02, Slide 015
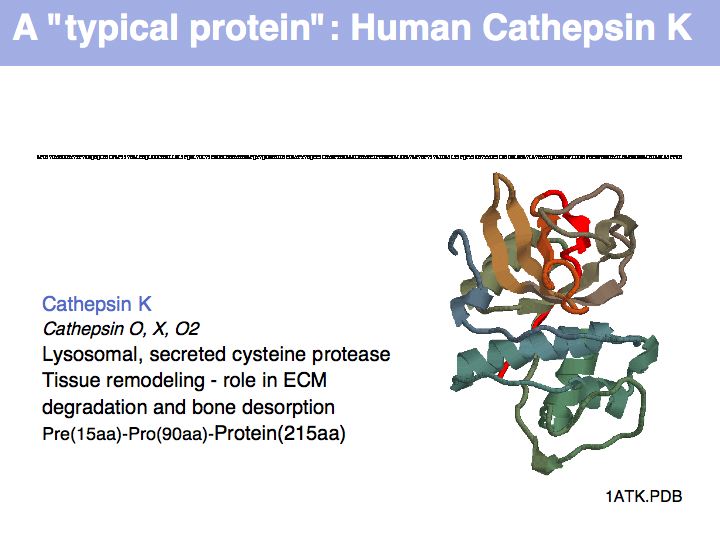
The physicochemical properties of amino acids determines their role in e.g. a folded protein structure. For example, consider the amino acid distribution in a typical enzyme, such as cathepsin K (1ATK).
The physicochemical properties of amino acids determines their role in e.g. a folded protein structure. For example, consider the amino acid distribution in a typical enzyme, such as cathepsin K (1ATK).
Slide 016

Lecture 02, Slide 016
Hydrophobic amino acids - the group FAMILYVW - are found predominantly in the core of a protein, small amino acids such as GASC are often found in turns, charged amino acid sidechains - (+):KRH and (-)DE - are almost exclusively found on the surface; the energetic requirements for desolvation of the sidechain makes their incorporation into the core unfavourable.
Hydrophobic amino acids - the group FAMILYVW - are found predominantly in the core of a protein, small amino acids such as GASC are often found in turns, charged amino acid sidechains - (+):KRH and (-)DE - are almost exclusively found on the surface; the energetic requirements for desolvation of the sidechain makes their incorporation into the core unfavourable.
Slide 017

Lecture 02, Slide 017
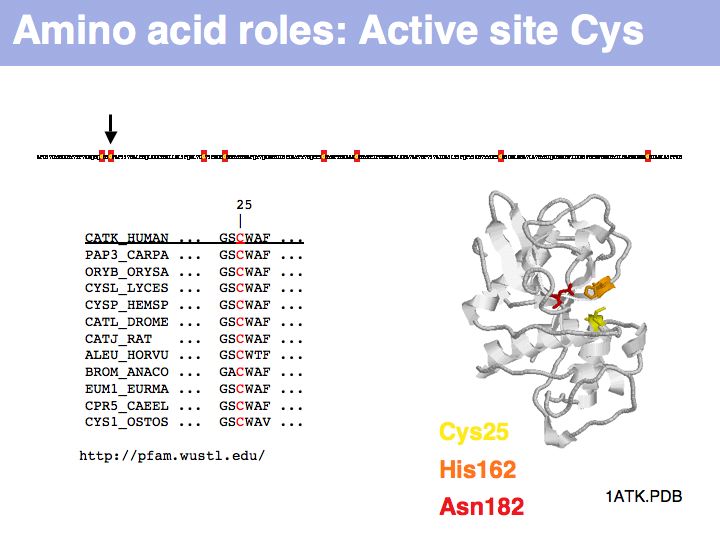
Cysteine can take on a number of different roles, depending on its context. Here, cysteine forms part of the active site, it is the nucleophile in the catalytic triad C-H-N; cathepsin is thuss an example of a cysteine-protease. This particular cysteine is absolutely conserved in related proteins.
Cysteine can take on a number of different roles, depending on its context. Here, cysteine forms part of the active site, it is the nucleophile in the catalytic triad C-H-N; cathepsin is thuss an example of a cysteine-protease. This particular cysteine is absolutely conserved in related proteins.
Slide 018

Lecture 02, Slide 018
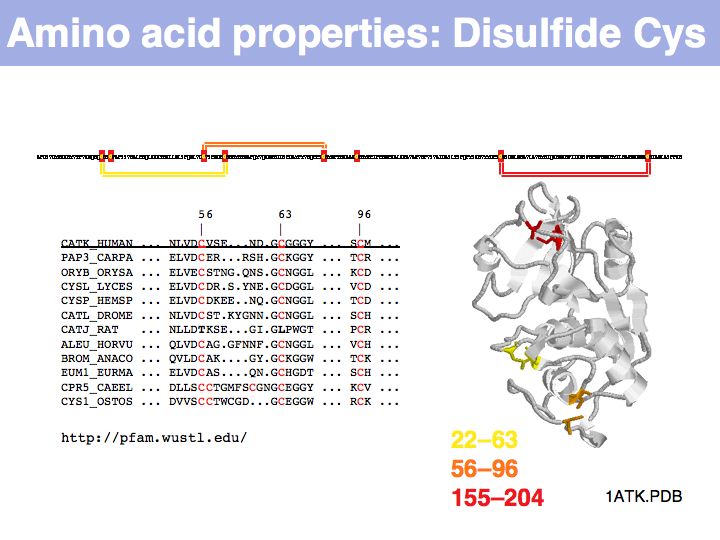
In secreted proteins only, cysteine often forms structural disulfide bridges in which two thiol groups oxidize to a covalent disulfide bond. These cysteines usually are highly conserved.
In secreted proteins only, cysteine often forms structural disulfide bridges in which two thiol groups oxidize to a covalent disulfide bond. These cysteines usually are highly conserved.
Slide 019

Lecture 02, Slide 019
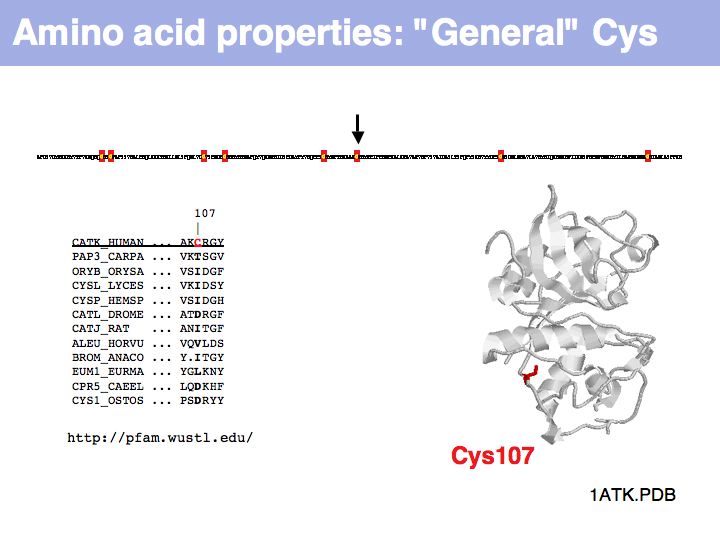
Cysteine can also be found in a very general role, simply as a somewhat polar, small residue. This general role is infrequent in secreted proteins since it can interfere with the formation of the correct disulfide topology and generally makes the protein sensitive to oxidation. Such cysteines are poorly conserved in related proteins.
Cysteine can also be found in a very general role, simply as a somewhat polar, small residue. This general role is infrequent in secreted proteins since it can interfere with the formation of the correct disulfide topology and generally makes the protein sensitive to oxidation. Such cysteines are poorly conserved in related proteins.
Slide 020

Lecture 02, Slide 020
Sequence is the most important abstraction in biology; you need to know your amino acids in order to relate a sequence back to the biopolymer. Required knowledge is: the structural formula, the one- and three- letter codes and key properties (such as charge, relative size, polarity) for all 20 proteinogenic amino acids.
Sequence is the most important abstraction in biology; you need to know your amino acids in order to relate a sequence back to the biopolymer. Required knowledge is: the structural formula, the one- and three- letter codes and key properties (such as charge, relative size, polarity) for all 20 proteinogenic amino acids.
Slide 021

Lecture 02, Slide 021
In Shakespeare's classic tragedy of romantic love and family allegiance, Juliet encapsulates the play's central struggle in this phrase by claiming that Romeo's family name is an artificial and meaningless convention. Just like in the world of the sequence abstraction, this is only partially true: the problems are not just based in the fact that Romeo is called a Montague, but that he is in fact a member of that family. Pure, abstract labels, of course, can be arbitrarily replaced.
In Shakespeare's classic tragedy of romantic love and family allegiance, Juliet encapsulates the play's central struggle in this phrase by claiming that Romeo's family name is an artificial and meaningless convention. Just like in the world of the sequence abstraction, this is only partially true: the problems are not just based in the fact that Romeo is called a Montague, but that he is in fact a member of that family. Pure, abstract labels, of course, can be arbitrarily replaced.
Slide 022

Lecture 02, Slide 022
Slide 023

Lecture 02, Slide 023
Slide 024

Lecture 02, Slide 024
Some ambiguity can only be resolved if the context of the representation is specified.
Some ambiguity can only be resolved if the context of the representation is specified.
Slide 025

Lecture 02, Slide 025
Slide 026

Lecture 02, Slide 026
Slide 027

Lecture 02, Slide 027
Often synonym constrained controlled vocabularies are presented as option lists on Web forms. If the CV is too long for this approach to be practical, defining the correct form becomes a challenge. In well engineered databases a lot of effort is spent on this task; typically a large dictionary of synonym mappings is employed in some form.
Often synonym constrained controlled vocabularies are presented as option lists on Web forms. If the CV is too long for this approach to be practical, defining the correct form becomes a challenge. In well engineered databases a lot of effort is spent on this task; typically a large dictionary of synonym mappings is employed in some form.
Slide 028

Lecture 02, Slide 028
NB. The situation that a unique property of an entity can be concisely described is the ideal case: in that case the identifier captures the most fundamental aspect of the molecule. For calcium, the element does not just have the atomic number 20, it is the element with 20 protons. Similarly oxytocin does not just have the sequence CYIQNCPLG, it is the peptide with that sequence. However, these are favourable exceptions and more commonly unique, abstract labels - such as identifiers - have to be defined.
NB. The situation that a unique property of an entity can be concisely described is the ideal case: in that case the identifier captures the most fundamental aspect of the molecule. For calcium, the element does not just have the atomic number 20, it is the element with 20 protons. Similarly oxytocin does not just have the sequence CYIQNCPLG, it is the peptide with that sequence. However, these are favourable exceptions and more commonly unique, abstract labels - such as identifiers - have to be defined.
Slide 029

Slide 030

Lecture 02, Slide 030
Slide 031

Lecture 02, Slide 031
Binary information vs ?strings?
Binary information vs ?strings?
Slide 032

Lecture 02, Slide 032
Slide 033

Lecture 02, Slide 033
Read more about the FASTA format.
Read more about the FASTA format.
Slide 034

Lecture 02, Slide 034
Slide 035

Lecture 02, Slide 035
Just like in human language, rigourous syntactical rules enforce that you can?t use bad grammar and get away with it.
Just like in human language, rigourous syntactical rules enforce that you can?t use bad grammar and get away with it.
Slide 036

Lecture 02, Slide 036
XML formatted files are human readable - in principle. The abundance of tags can make this challenging in practice. The loss of readability is a trade-off for the gain in rigour.
XML formatted files are human readable - in principle. The abundance of tags can make this challenging in practice. The loss of readability is a trade-off for the gain in rigour.
Slide 037

Lecture 02, Slide 037
Slide 038

Lecture 02, Slide 038
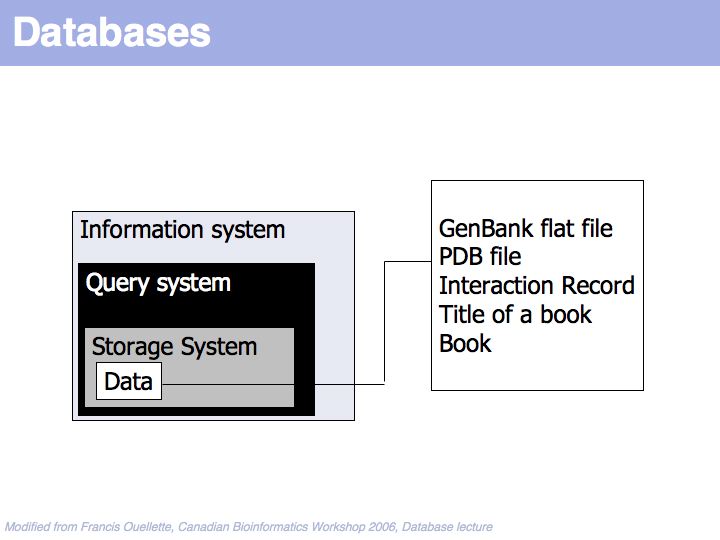
A "Database" is much more than just the data it contains.
A "Database" is much more than just the data it contains.
Slide 039

Lecture 02, Slide 039
At the core of the database is the data and of course the data should be the primary focus of attention of the database providers. Issues are the correctness of the data, mechanisms to validate and maintain records, frequency of updates, consistency etc.
At the core of the database is the data and of course the data should be the primary focus of attention of the database providers. Issues are the correctness of the data, mechanisms to validate and maintain records, frequency of updates, consistency etc.
Slide 040

Lecture 02, Slide 040
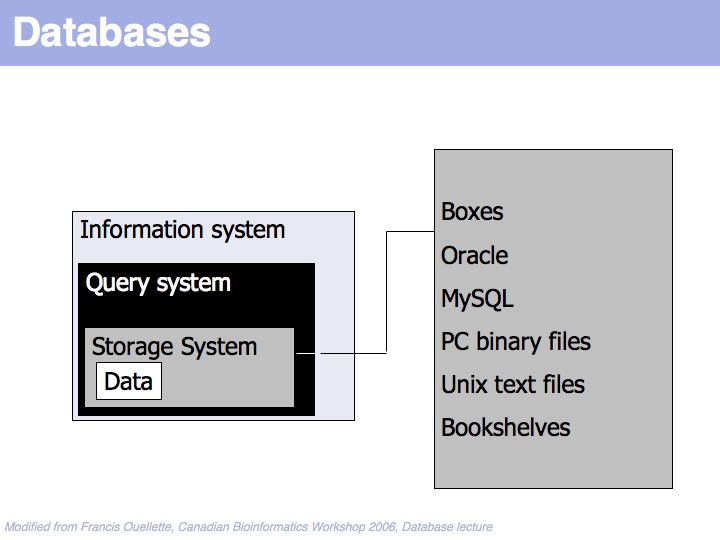
Data is stored in some storage system.
Data is stored in some storage system.
Slide 041

Lecture 02, Slide 041
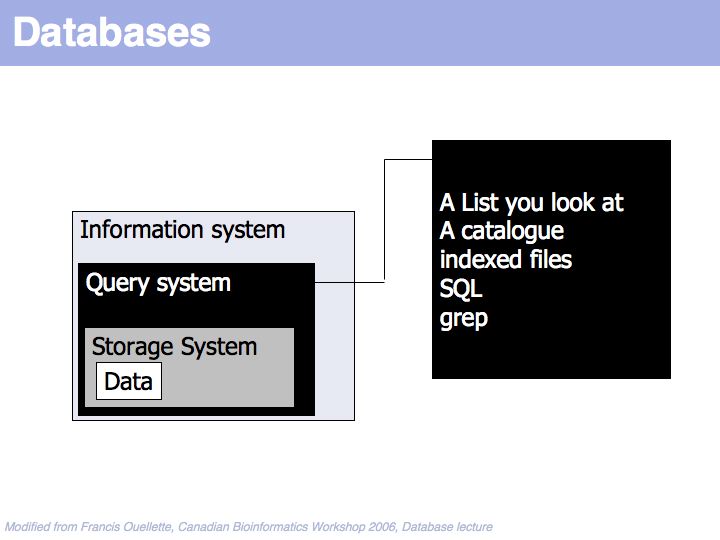
Data is found through some query system.
Data is found through some query system.
Slide 042

Lecture 02, Slide 042
Storage resources, query and maintenance functions, and meta-information services are all united into a common interface that is presented to the user.
Storage resources, query and maintenance functions, and meta-information services are all united into a common interface that is presented to the user.
Slide 043

Lecture 02, Slide 043
Slide 044

Lecture 02, Slide 044
Slide 045

Lecture 02, Slide 045
Slide 046

Lecture 02, Slide 046
Slide 047

Lecture 02, Slide 047
Slide 048

Lecture 02, Slide 048
Slide 049

Lecture 02, Slide 049
Slide 050

Lecture 02, Slide 050
Slide 051

Lecture 02, Slide 051
Slide 052

Lecture 02, Slide 052
Slide 053

Lecture 02, Slide 053
Slide 054

Lecture 02, Slide 054
Slide 055

Lecture 02, Slide 055
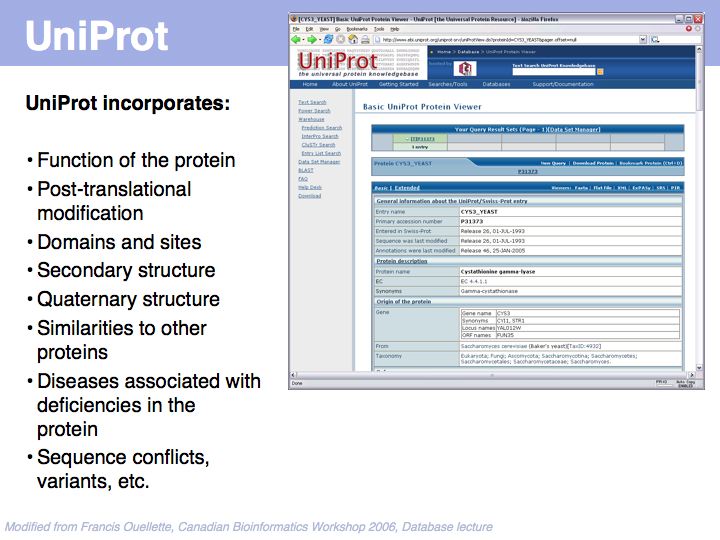
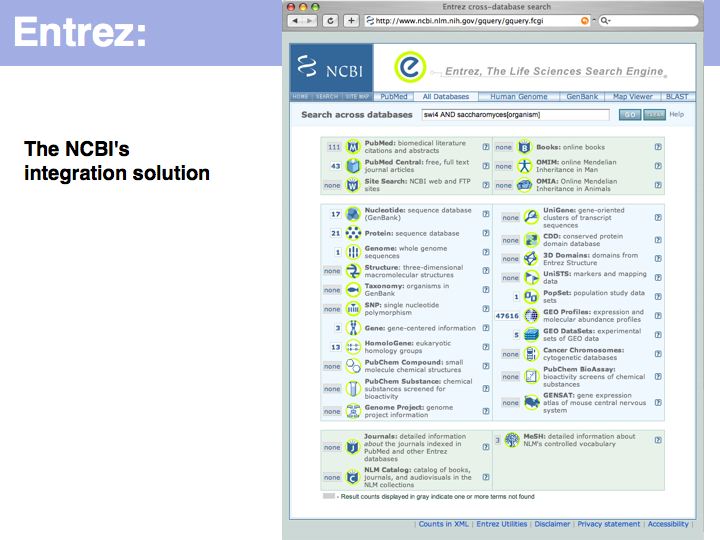
UniProt is arguably the more comprehensive resource, however it is not integrated with the GenBank world, although that would be reasonably straightforward to do. For example, neither does the NCBI record for the Swi4 protein contain a reference to the UniProt accession number, nor does the UniProt record for the same protein contain an NCBI accession number or a GI. National database politics are not always in the best interest of the worldwide scientific community.
UniProt is arguably the more comprehensive resource, however it is not integrated with the GenBank world, although that would be reasonably straightforward to do. For example, neither does the NCBI record for the Swi4 protein contain a reference to the UniProt accession number, nor does the UniProt record for the same protein contain an NCBI accession number or a GI. National database politics are not always in the best interest of the worldwide scientific community.
Slide 056

Lecture 02, Slide 056
Slide 057

Lecture 02, Slide 057
SwissProt records - a subset of the UniProt Knowledge Base - are the highest standard, manually curated, non-redundant protein records available. Unfortunately the growth of the sequence databases far exceeds any human curation capabilities!
SwissProt records - a subset of the UniProt Knowledge Base - are the highest standard, manually curated, non-redundant protein records available. Unfortunately the growth of the sequence databases far exceeds any human curation capabilities!
Slide 058

Lecture 02, Slide 058
Slide 059

Lecture 02, Slide 059
Slide 060

Lecture 02, Slide 060
Slide 061

Lecture 02, Slide 061
Slide 062

Lecture 02, Slide 062
Slide 063

Lecture 02, Slide 063